Spinal motor nöron, periferik afferent liflerin, interneronların ve supraspinal yapılardan inen yolların kalıcı etkisi altındadır. Spastisite, monosinaptik refleksin abartılmasına karşılık gelir (Şekil 3.1). Merkezi sinir sisteminin çeşitli lezyonları, özellikle supraspinal inen yolların kontrollerini ve özellikle retikülospinal yolları etkiliyorsa, spastisiteye neden olur.
Spastisite, belirli bir uzuvdaki farklı kas grupları arasında farklı bir dağılım gösteren, motor yolların lezyonundan sonra meydana gelen değişken gecikmelerle dışsal, propriyoseptif veya interoseptif fizyolojik uyaranlara farklı tepkiler ile kendini gösteren klinik çeşitliliğe sahiptir. Sebep olan lezyonların etiyolojisi farklıdır ve lezyon yerleri çok değişkendir.
Bu klinik değişkenliğe karşılık olarak, çok çeşitli deneysel modeller vardır: beyin sapının Sherrington tarafından interkoliküler seviyede transeksiyonu, anında deserebrasyona sebep olur; tam spinal kord kesilme modeli; motor kortikal bölgelerinin (4 ve 6. alan) rezeksiyon modeli; son olarak, spinal iskemi modeli, motor nöronal hiperaktivite ile interneuronal yoksunluğa neden olur.
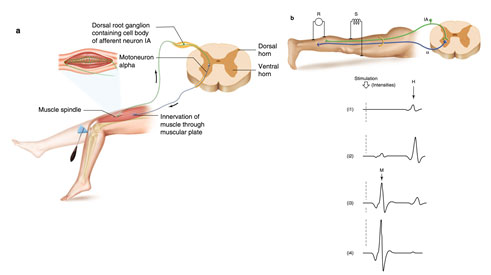
3.1 (a) Miyotatik refleks. Miyotik refleks, gerilmesine yanıt olarak bir kasın kasılmasıdır (örneğin, patellar tendon bir tendon refleks çekici ile dokunduğunda). Bu refleks kas uzunluğunun otomatik olarak düzenlenmesini sağlar. Bir kas uzatıldığında, duyusal reseptörü olan mil gerilir ve sinir aktivitesini (IA lifi) arttırır. IA'daki afferent uyarı serisi alfa motor nöronu uyarır, kasın kasılmasına neden olur ve böylece gerilmeye karşı direnir. (b) H refleksi. Monosinaptik yolları (IA afferent fiber ile alfa motor nöron) inceleyen H refleksini kaydetme yöntemi. Popliteal fossada tibial sinirin uyarılması (S) ve soleus kasının motor yanıtının kaydedilmesi (R). Stimülasyon yoğunluklarının arttırılması H refleksinin (I1, I2) ortaya çıkmasına neden olur. Stimülasyon yoğunlukları artmaya devam ettiğinde, H refleksi, doğrudan motor yanıtının artan genliği (M; I3, I4) ile ortaya çıkan motor liflerinin toplanmasıyla ile kaybolur.
3.1 Spinal Kordun Organizasyonu
3.1.1 Alpha Motor Nöronlar
Alfa motor nöronlar ventral boynuzda bulunur. Aynı kas grubunu innerve edenler, 2-4 metamere uzatabilen uzunlamasına motor sütunlarında organize edilmiş bir motor nöron grubuyla ilişkilidir. Spinal alfa motor nöronları, Şekil 3.2'de şematik olarak özetlendiği gibi somatotopik bir dağılıma sahiptir.
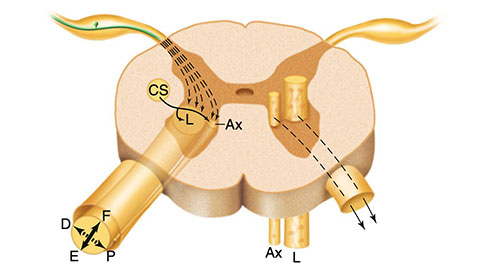
Şekil 3.2 Ventral boynuzdaki motor nöronlar, hedeflerine ve fonksiyonel rollerine göre somatotopik olarak iki boyuna sütun halinde düzenlenir: medial grup (medial sütun) aksiyel kasları (boyun ve gövde) ve lateral grubu (lateral sütun), ekstremite kaslarını (üst ekstremite ve alt ekstremite) innerve eder. Bu grupta ekstansör kaslar için motor nöronlar daha ventral olarak yer alırken, fleksör kaslar için motonöronlar daha dorsal olarak yerleşmiştir. Ax aksiyal kas sistemi, L uzuvlar, D uzuvların distal kasları, P uzuvların proksimal kasları, F fleksör kaslar, E ekstansör kaslar, CS kortikospinal sistem
3.1.2 Primer Afferentler
Rexed'in tüm laminaları, interkomissüral değişim alanı olan lamina VIII hariç, doğrudan primer afferentleri alır. Büyük çaplı lifler lamina III, IV, V, VI ve IX üzerine, küçük çaplı lifler yüzeysel lamina I ve II'ye ve daha derin lamina V ve X üzerine yayılım yapar. Bazı propriyoseptif IA lifleri monosinaptik refleksi tetiklemek için doğrudan lamina IX motor nöronlar üzerine yansır. Bununla birlikte, bu propriyoseptif afferentlerin çoğunluğu dorsal boynuzun tabanındaki lamina VI’da biter, bu da primer afferent liflerin çıkan yolların kaynağı olan internöronlarla bağlandığı gerçek bir “propriyoseptif tabaka” oluşturur.
Dorsal köklerin giriş bölgesi için insanlarda anatomik ve fizyolojik çalışmalar, birincil afferentlerin boyutlarına ve omurilikteki hedeflerine ve dolayısıyla işlevlerine göre uzamsal bir organizasyonun varlığını ortaya koymuştur. Bu nedenle, dorsal kök giriş bölgesinin ventrolateral bölgesinde, esas olarak nosiseptif olan ince lifler (Aδ ve C) yeniden gruplanır (bakınız Bölüm 9); daha sonra, dorsal boynuzunun en üst katmanlarına ulaşmak için Lissauer yolunun medial kısmından geçerler. Dorsal kök giriş bölgesinde dorsomedial olarak yer alan büyük lifler (Aβ), dorsal kolona ulaşır. İşlevleri, epikritik dokunsal ve bilinçli propriyosepsiyon türleri hakkında bilgileri beyin sapındaki lemniskal yollara taşımaktır.
Affentler tarafından salınan nörotransmitterler esas olarak uyarıcı amino asitlerdir (glutamat, aspartat). Sadece yüzeysel tabaka I ve II üzerine giren küçük çaplı afferentler tarafından salınan nörotransmiterler inhibe edicidir (γ-aminobutirik asit [GABA], glisin). Lamina I nöronları sadece glisinerjik sinaptik inhibisyon alırken, lamina II nöronlarının yaklaşık yarısı saf GABAerjik ve geri kalanı karışık GABAerjik ve glisinerjik inhibisyon alır.
3.1.3 Spinal İnterneron Sistemi
Propriyospinal interneronlar adı verilen omuriliğin internöronları üç tiptir. Çoğu kısa sinir lifine sahiptir; bazıları orta ve bazıları uzun sinir liflerine sahiptir. Hücre gövdeleri gri maddenin tüm tabakasında bulunur. Bununla birlikte, ara bölge (VII ve VIII lamina) içinde yer alan internöronlar özellikle bölgeler arası spinal refleksleri kontrol eder. Kısa internöronların aksonları, ağırlıklı olarak lamina IX'un lateral kısmındaki fleksör kaslarının motor nöronlarına iletmek için 4-8 komşu metamerde dorso ve ventrolateral yollara gider. Ara internöronlar, aksonlarını, daha da büyük komşu metamerlerin lamina IX'ındaki, özellikle ekstansör kasların motor nöronlarının kolonlarındaki ventrolateral yoldan gönderirler.
Bu internöronların hepsi, bilgiyi sadece periferal orijinden değil, aynı zamanda suprasegmental iniş yollarından getiren liflerin hedefleridir. Bu bilgileri işledikten sonra, internöronlar, motor havuzları üzerinde engelleyici veya kolaylaştırıcı bir uyarı komutu gönderir. Fizyolojik olarak, çeşitli interneronlar vardır. IB afferentleri tarafından aktive edilen otojenik inhibisyon refleksinin nöronları, lamina V ve VI'nın lateral kısmında bulunur. Refleks resiprokal IA liflerini destekleyen interneronlar, lamina VII'de bulunur. Fleksiyon reflekslerinde yer alan interneronlar, lamina VII'nin dorsolateral kısmında bulunur. Öte yandan, alfa motor nöronlarının tekrarlayan inhibisyonunun internöronları olan Renshaw hücreleri, lamina VII'nin en ventral kısmında bulunur.
İnhibitör internöronların baskın nörotransmitterleri olarak glisin vardır. Diğer internöronların çoğu GABAerjik ve enkefalinerjiktir. Renshaw hücreleri esas olarak glisinerjik ve bazı durumlarda GABAerjiktir.
Asetilkolin kullanan rekürren motor nöron lifleri tarafından aktive edilirler. Aynı alfa veya gama motor nöronları üzerinde inhibe edici (hiper-polarize) bir etkiye sahiptirler. Bu Renshaw internöronları, afferent lifler ve supraspinal kaynaklı inen yollar tarafından modüle edilir. Bir başka GABAerjik internöron popülasyonu, afferent liflerin presinaptik, aksono-aksonal inhibisyonu yoluyla etki eder. Reseptörler, GABA-A (bir transmembran Cl kanal benzodiazepin agonisti ile birleştirilmiş) veya GABA-B (K + kanalı baklofen agonisti) olabilir. Aktive edilmiş GABA-A ve GABA-B reseptörleri membran hiperpolarizasyonuna yol açar ve böylece motor nöron uyarılabilirliğini azaltır. Bu inhibitör internöronlar, primer afferentler veya inen monoaminerjik yolaklar yoluyla aktive edilebilir. Bunlar alfa-2 (noradrenerjik yollar için) ve 5-HT1 (serotonerjik yollar için) reseptörlerini aktive eder. Enkefalinerjik interneronlar, dorsal boynuzun yüzeysel tabakasında çok sayıda bulunur ve özellikle opioid reseptörleri mu, delta ve kappa yoluyla nosiseptif transmisyon üzerinde inhibitör bir etki gösterir.
3.1.4 Çıkan Yollar
Spinoserebellar traktus esasen Clarke ve Betcherew çekirdeklerinin lamina VI’sı ile lamina IV ve IX'un aksonlardan oluşan ikincil nöronlardır. Bu çıkan yollar lateral sütunda seyahat eder ve uzuvlardan (Gowers fasculususu) bilgi taşıyan propriyoseptif kontralateral liflerin ventral demetine (Flechsig fasciculus'u) ve gövdeden bilgi taşıyan dorsal demete ayrılır. Ventral kolon, ipsi ve kontralateral yolaklar ile VII, VIII ve X tabakalarının derin katmanlarından kaynaklanan spinoretiküler yolu barındırır. Bu sistem sadece nosisepsiyonda değil, aynı zamanda retiküler oluşum yoluyla motor kontrol alanında da rol oynar.
3.2 Supraspinal İnen Yollar
3.2.1 Kortikospinal Sistem
Frontoparietal korteksin geniş alanlarından (ör. Brodmann alanları 4, 3a, 3b, 1, 2, 5,
SII, supplamenter motor alanı) kaynaklanan lifler, kortikospinal lifler olarak adlandırılır ve bu aksonlar kortikal bölgelerin iç katmanı olan V katmandaki internal piramid tabakası denilen yerdeki nöronların aksonlarını oluşturur.
Kortikospinal liflerin çoğu medüller piramitte çaprazlaşır (dekussasyo pramidarium) ve lateral kolonun medial kısmından inerler. Ventral boynuzdaki alfa motor nöronlarına yayılırlar. Az miktarda lif ipsilateral kalır ve motor nöronlara metamerik projeksiyon seviyesinde çaprazlanıp geçmeden önce ventral kolondan iner. Bu kortikospinal liflerin çoğu ventral boynuzda sonlanır, ancak bazıları da dorsal boynuz ve ara bölge üzerine yayılan lamina IV ila IX üzerinde son bulur. Parietal orijinli lifler tercihen dorsal boynuz (laminae IV-VI) üzerinde yayılırken, motor korteksten lifler tercihen ara bölgenin interneronları ve ventral boynuz üzerinde yayılır.
3.2.2 İnen Lateral Sistem
İnen lateral sistem, lateral pontin ve medüller retiküler formasyondan (giganto-hücresel çekirdek) gelen rubrospinal ve retikülospinal yolları içerir. Bu yollar dorsolateral kolonda yeniden gruplandırılır ve karşı tarafa geçerek lamina I ve lamina V, VI, VII ve IX'un lateral kısmına yayılır.
3.3 Retiküler Formasyonun Önemi
Aşırı spastisite ile etkilenen hastaların çoğu, özellikle de serebral palsi olan çocuklar, özürlülüklerini güçlü bir şekilde şiddetlendiren aşırı reaktiviteye bağlı ek belirtilerle daha kötü duruma girerler. Bu klinik, diğerlerinin yanı sıra, küçük fiziksel veya psikolojik irritasyonlarla bile parazit motor reaksiyonlarının, herhangi bir yakın veya uzak uyarana karşı uyumsuz hareketlerin oluşturduğu büyük tepkiler ile kendini gösterir.
Tüm merkezi sinir sistemi bu kliniğe sadece propriyospinal internöron ağı üzerinden değil, aynı zamanda önemli ölçüde retiküler formasyon (RF) ve yaygın bağlantıları yoluyla da dahil olur.
RF, tüm beyin sapı tegmentumunda gevşek bir şekilde nükleer gruplara yerleştirilmiş geniş aralıklı nöronlardan oluşur. Dev dendritik ağaçlar ve çok dallı afferent ve efferent akson ağı ile RF perikarya’sını diğerlerinden ayırıp, mikroskobik kesitlerde “retiküler” görünüme neden olur.
RF, tüm tegmentum boyunca sürekli olarak ilerler. Kranial sinir çekirdeği, ek duyu ve motor çekirdeği ve uzun ve kısa yollar tarafından işgal edilmeyen tüm alanı doldurur. Medulloservikal bileşke genellikle kaudal limiti olarak kabul edilir. Anatomik bakış açısından, RF'nin rostral sınırı orta beyin-diensefalon birleşimindedir, ancak fonksiyonel olarak omurilik ile diensefalik ve serebral bağlantılar arasındaki bir sürekliliği temsil eder.
RF, merkezi sinir sisteminin en heterojen bağlantılarına sahiptir. Epigramatik olarak ifade edilen RF, tüm merkezi sinir sisteminden impulslar alır ve ona impuls gönderir; böylece tüm zihinsel, motor ve duyusal fonksiyonları etkiler. Fizyolojik olarak RF nöronları, takviye etme, uyuşmazlık ve sonradan deşarj ile karakterizedir. RF, lemniskal ve diğer spesifik afferent duyu sistemleri üzerinde çok daha az etkisi olan genel anesteziklere ve bazı psikoaktif ilaçlara özellikle duyarlıdır.
3.3.1 RF Afferentleri
RF, bir nöral yolun kökenine, omurilikte veya beyinde olmasına ve yolun visseral veya somatik, motor veya duyusal fonksiyonunda olmasına bakılmaksızın, beyin sapı boyunca hem çıkan hem de inen yollardan kollateraller alır.
Afferentler lateral, parviselüler çekirdeğinde sinaps oluşturma eğilimindedir, bunlar daha sonra RF efferentlerini dağıtan gigantocellular medial çekirdeğe aktarılır. Bir istisna medial lemniscus'dur. Dorsal kolonlardan diskrimitif duyuları aktaran lifleri görünüşte RF'ye dallar göndermez; ancak çıkışından sonra medial lemniscus'a katılan lateral spinotalamik ve trigeminotalamik yollar RF'ye dallar gönderir. Ayrıca spinoretiküler traktuslar RF'ye kuvvetle yayılır.
RF'yi baypas eden afferent fiberler, iletilen bilgilerin topografik sırasını korumak için tasarlanmış ayırt edici veya birebir sistemlerdir. Bunlar dorsal kolon yolunda uzanan medial lemniscus, retino-genikulo-kalkarin traktus ve işitsel sistemin tonotopik liflerini içerir. İşitsel ve optik yollar RF'ye, yüksek sese veya ani parlak ışığa karşı hızlı bir şekilde tepki vermesinden anlaşıldığı gibi ulaşır, ancak bu sistemlerin bire bir bileşeni RF'yi atlar.
3.3.2 Spesifik İnen RF Yolları
İnen yolaklar retikülobulbar ve retikülospinal yollar ve spesifik nörotransmitter yolaklarıdır. Retikülobulbar yollar tüm kranial sinir çekirdeklerine doğru ilerler. Retikülospinal yollar omuriliğin ventrolateral kolonları içinde uzanır. Bu yollar medial RF çekirdeklerinden kaynaklanır ve pontin ve medüller retikülospinal yolları içerir.
Pontin retikülospinal yollar pontin retiküler çekirdeklerden kaynaklanır. Medial longititünal fasikülüsün inişinden sonra, tüm spinal kord seviyelerinde VII ve VIII tabakasında biterler ve vestibüler çekirdeklerin sonlandırma alanlarıyla geniş ölçüde örtüşürler. Bu yollar, postural reflekslere ve kas tonusuna aracılık etmek için aksiyel kaslara, özellikle de boynun kaslarına etki eder.
Medüller retikülospinal traktus nükleus retikularis gigantocellularis'ten ve VII ve IX tabakadaki tüm spinal kord seviyelerindeki sinapslardan kaynaklanır. Sonlandırma alanları, kortikospinal ve rubrospinal kanalların terminalleri ile çakışır.
Retikülospinal aksonların az bir kısmı ventral motor nöronlarda, çoğu VIII ile IX arasındaki laminaların internunsial nöronlarında sona ermektedir.
Sonuç
Spastisite, miyotatik refleksin abartılmasına karşılık gelir ve spinal hipereksitabilite ile bağlantılıdır. Bu hipereksitabilite, supraspinal inen yollardaki inhibitör kontrolün kaybından kaynaklanır (Şekil 3.3).
Spastisite, bu bölümde özetlendiği gibi, motor sistem fonksiyonundaki geniş ve çeşitli bozukluğun bir parçasıdır (Şekil 3.4).
Patolojik süreçten sonra ortaya çıkan spinal plastisite de rol oynar. Aşağıdaki iki mekanizmaya göre özetlenebilir: lezyonlu sinir liflerinin dejenerasyonu ile boşalan sinaptik bölgeler yönünde aksiyal büyüme fenomenleri (kollateral filizlenme (sprouting)) ve daha az ölçüde, presinaptik afferentlerin kaybını telafi eden nöronal reseptörlerin aşırı duyarlılık fenomeni.
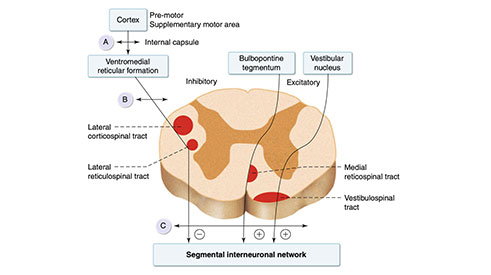
Şekil 3.3 Supraspinal kontrol uygulayan büyük inen sistemler. Anatomik ilişkiler ve inhibitör ve uyarıcı inen sistemler arasındaki kortikal kontrol açısından farklılıklar, üst motor nöron lezyonunun anatomik konumunun, ortaya çıkan klinik paternin belirlenmesinde önemli bir rol oynadığını gösterir. A: Ana inhibitör sistemin, yani lateral retikülospinal sistemin yapılarını kolaylaştıran kortikospinal sistemi ve kortikoretiküler yolu etkileyen lezyon. B: Kortikospinal lifleri ve lateral retikülospinal yolu etkileyen inkomplet omurilik lezyonu. C: Lateral retikülospinal lifler ve uyarıcı yolaklar, yani medial retikülospinal kanal ve vestibulospinal kanal ile birlikte tüm kortikospinal lifleri etkileyen komplet omurilik lezyonu,. Artı sembolü uyarıcı yolu gösterir; eksi simgesi inhibitör yolu gösterir. Uyarıcı yolların fleksör refleksler üzerinde inhibitör etkileri vardır.
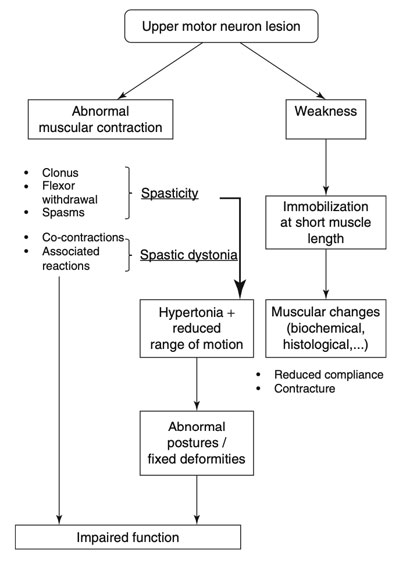
Şekil 3.4 Üst motor nöron lezyonunda fonksiyon bozukluğuna yol açan patofizyolojinin şematik gösterimi (Sheean G (2008) Spastisite nörofizyolojisi. In: Barnes MP, Johnson GR (eds) Üst motor nöron sendromu ve spastisite - klinik yönetim ve nörofizyoloji, 2. edn. Cambridge University Press, Cambridge, s 27)